Kv10.2
Description: potassium voltage-gated channel, subfamily H (eag-related), member 5 Gene: Kcnh5 Alias: Kv10.2, eag2, h-eag2, kcnh5
Kv10.2, encoded by the KCNH5 gene, is a potassium channel, voltage-gated, subfamily H. Kv10.2 is natively expressed in the nervous system [1709]
Experimental data
Rat Kv10.2 gene in CHO host cells datasheet |
||
|
Click for details 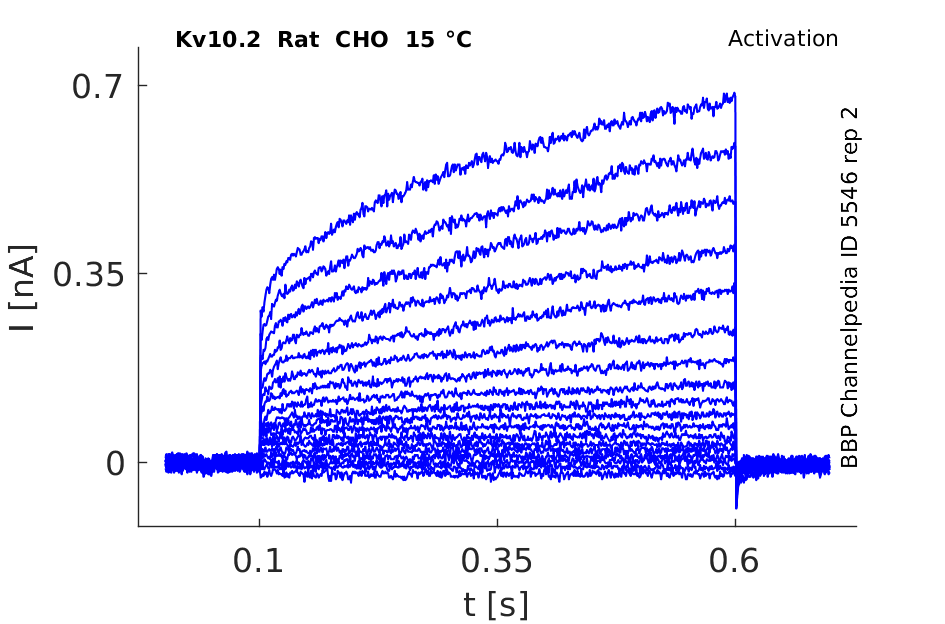
15 °Cshow 91 cells |
Click for details 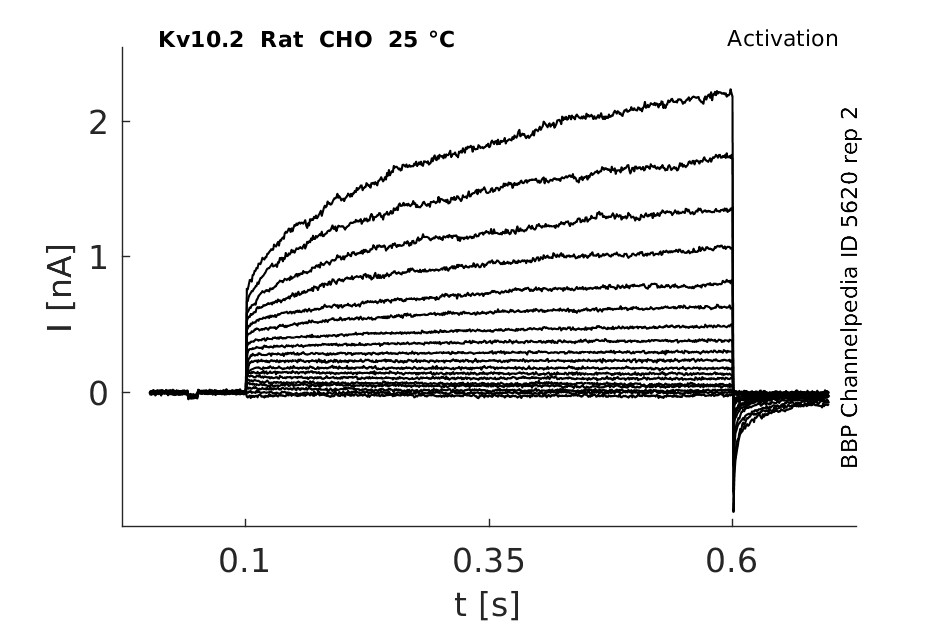
25 °Cshow 76 cells |
Click for details 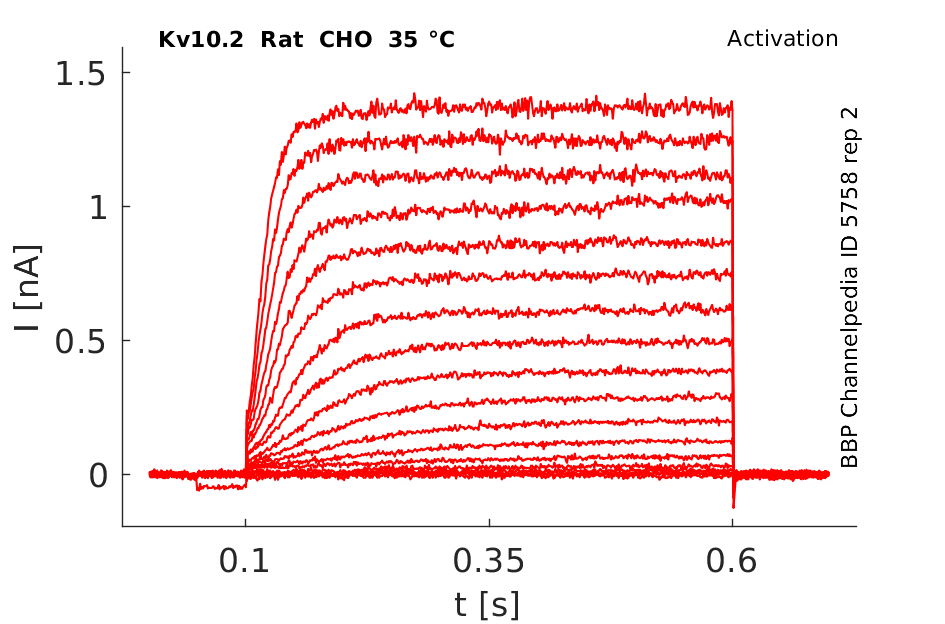
35 °Cshow 104 cells |
Kv10.2 belongs to the ether-à-go-go (EAG) family within the voltage-gated potassium (Kv) channel superfamily.
Transcript
| Species | NCBI accession | Length (nt) | |
|---|---|---|---|
| Human | NM_139318.5 | 11283 | |
| Mouse | NM_172805.3 | 4061 | |
| Rat | NM_133610.2 | 3505 |
Protein Isoforms
Isoforms
Post-Translational Modifications
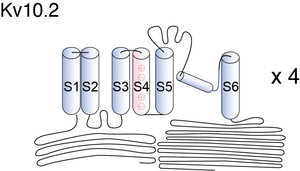
Visual Representation of Kv10.2 Structure
Methodology for visual representation of structure available here
Structural modelling of Kv10.2 Arg327 in six states

Kv channels are membrane proteins that consist of six transmembrane segments, including S1–S4 as the VSD, a relative independent domain that transforms membrane potential changes into channel gating, and S5 and S6 as the pore-forming module. Sequence analysis shows that Arg327 is located in the critical S4 helix of the VSD (Fig. 1A). Within the S4 helix of VSD, Arg327, together with Lys325, Arg329, and Arg331 (commonly known as R[K]1–R4), are positively charged residues that sense changes in the membrane potential and produce a conformational change of the VSD, which in turn controls channel gating [1709]
Similar to other Kv channels, the functional Eag channel is assumed to be a tetramer, each monomer consisting of six putative transmembrane domains, with the S4 domain serving as the main voltage sensor. In addition, they also possess long intracellular N- and C-termini [778], [640]. The binding site for divalent cations requires acidic residues in S2 and S3 that are unique to the EAG family [776]. The homologous residue Serine 321 in Kv10.2 contributes to effects of Mg2+ and Ni2+ [770].
Not only transmembrane spanning regions, but also non-membrane spanning regions are responsible for differences in the behaviour of the hEag1 and hEag2 potassium channels. [749]
Kv10.2 predicted AlphaFold size
Methodology for AlphaFold size prediction and disclaimer are available here
Electrophysiology of rat eag2

Heag1 and heag2 comparison

In the Xenopus oocyte expression system the time course of activation was about four times slower and the voltage required for half-maximal subunit activation was about 10 mV greater for hEAG2 channels. However, its voltage dependence was smaller and, therefore, hEAG2 channels start to open at more negative voltages than hEAG1. Coexpression of both isoforms and kinetic analysis of the resulting currents indicated that they can form heteromeric channel complexes in which the slow activation phenotype of hEAG2 is dominant [1713]
Heag1 and Heag2 in X oocytes

Factors effecting Kinetics
Eag channels, eag1 and eag2, are characterized by outward rectification without inactivation, and by fast deactivation and slow activation. The activation kinetics depend on the holding potential (‘‘Cole–Moore shift’’) and extracellular magnesium [81]
Markov Model

Markov modelling demonstrated that simple 5- and 6-state models were inadequate and that a 10- or 12-state model was more appropriate for description of EAG2 channels [1504]
Hodgkin and Huxley Model for rEAG

Kv10.2 expression in brain
The rat eag2 mRNA was predominantly expressed in neural tissue and was not detected in adult skeletal, cardiac, or smooth muscle. Within the brain, its distribution overlaps that of rat ether-a` -go-go1 in specific regions within the cortex and olfactory bulb, but was differentially distributed in other locations, being scanty within the cerebellum, and most notably present in the thalamus, inferior colliculus, and certain brainstem nuclei [800]
In the central nervous system, Eag2 levels are the most prominent in layer IV of the cerebral cortex, an area for which Eag2 has been proposed to participate in the regulation of the behavioral-state-dependent entry of sensory information (Saganich et al.,1999). Similar immunoreactivity patterns for Eag1 and Eag2 are observed in the granular layer of the dentate gyrus and pyramidal layers of the CA1, CA2, and CA3 subfields of the hippocampus [1710]
Kv10.2 expression in body
The human genome has two isoforms of Eag channels, hEag1 (KCNH1, Kv10.1) and hEag2 (KCNH5, Kv10.2). Both are expressed mainly in the central nervous system, although hEag2 has also been found in skeletal muscle, heart, lung, liver, placenta, kidney and pancreas [779], [777].
Distribution of Kv10.2 on neuron
Both Kv10.1 and 10.2 channels are expressed largely in neurons and localized at synapses [1711]
Distribution of Eag1 and Eag2 in Hippocampal neurons
Immunofluorescence studies using markers for different compartments of neurons demonstrated a differential expression pattern of rEag1 and rEag2 K+ channels in the somatodendritic region. Furthermore, rEag1 K+ channels were in close proximity to synaptophysin and densin-180, but not GAD65. Our data suggest that both rEag1 and rEag2 K+ channels may play a pivotal role in the regulation of the excitability of dendrites and somas, and that rEag1 K+ channels may modulate the postsynaptic signaling of glutamatergic synapses [1714]
Function in CNS
Although the functional role of Eag1, Eag2, and SK3 in the central nervous system remains undetermined, Eag2 mRNA downregulation might be related to hippocampal impairment of learning and memory [1710]
Anxiety
The hippocampus has a well-established role in spatial learning and memory, and it may also regulate defensive behaviors related to anxiety. Recently, it was demonstrated that isolation-reared rats had reduced Eag2 mRNA expression in the hippocampus, prefrontal cortex, and striatum. This finding was related to a decrease in exploratory activity in a novel environment, suggesting the involvement of Eag2 in stress-related responses [1710]
Expoloratory action
solated and socially reared rats were tested for exploratory activity in the open field test for 5 min and brains were processed for reverse-transcription coupled to quantitative polymerase chain reaction (qRT-PCR). Isolated reared rats showed decreased exploratory activity in the open field. Compared to socially reared rats, isolated rats showed reduced Htr2a mRNA expression in the striatum and brainstem and reduced Eag2 mRNA expression in all examined regions except cerebellum.
Tumor Growth
Medulloblastoma (MB) is the most common pediatric CNS malignancy. We identify EAG2 as an overexpressed potassium channel in MBs across different molecular and histological subgroups. EAG2 knockdown not only impairs MB cell growth in vitro, but also reduces tumor burden in vivo and enhances survival in xenograft studies. Therefore voltage-gated potassium channel EAG2 controls mitotic entry and tumor growth in medulloblastoma via regulating cell volume dynamics [1715]
Neurological disease
Voltage-gated potassium channel Kv10.2 (KCNH5) is expressed in the nervous system, but its functions and involvement in human disease are poorly understood. We studied a human Kv10.2 channel mutation (R327H) recently identified in a child with epileptic encephalopathy and autistic features [1709]
Mg,Ni
Kv10 channel gating is that the activation rate is highly sensitive to divalent cations such as Mg2+ and Ni2+ [774],[775]. These ions preferentially slow the first independent transition and thus bind a closed conformation early in the activation pathway [772], [773]. The binding site for divalent cations requires acidic residues in S2 and S3 that are unique to the EAG family [776]. Silverman et al. [776] proposed that divalent cations electrostatically slow movement of the third S4 gating charge into an extracellular pocket and Schonherr et al. [777] proposed that Mg2+ slows activation sterically by narrowing the gating canal.
R327H mutation
Recently, we identified a human Kv10.2 channel mutation (R327H) in a child with epileptic encephalopathy with seizures intractable to medical therapy and autistic features via exome sequencing. To understand how this R327H mutation, which substitutes a single residue within the voltage-sensing helix S4, affects the structure and function of Kv10.2, we performed this structural and functional study. [709]
Calcium
Whole cell currents under our standard recording conditions (with 20 nM estimated free calcium in the pipette medium) showed a tendency to decline, with or without 2 mM ATP in the patch pipette. From initial current magnitudes observed at [1711]
Quindine
Upon expression in mammalian cells, quinidine blocked hEAG1 channels (IC50 1.4 WM) more potently than hEAG2 channels (IC50 152 WM), thus providing a useful tool for the functional distinction between hEAG1 and hEAG2 potassium channels [1713]
Mutation at I345I residue
To investigate whether this residue plays a role in determining differ- ences in activation between heag1 and heag2 channels, we mutated the residue, I345L in heag1, and L345I in heag2. In neither case, though, did the mutation affect the activation kinetics: the heag1 mutant remained fast and the heag2 mutant remained slow. This therefore shows that this residue, and indeed the whole of the S4–S5 linker, do not participate in determining differences in activation kinetics between the two channels [81]
Cytoskeletal Elements
hEAG channels establish complex interactions with cytoskeletal elements, and that these interactions strong- ly influence the properties of the channels. We also con- clude that the electrophysiological changes observed during the cell cycle are most likely due to reorganiza- tion of the cytoskeleton during the G2/M transition
References
Wray D
The roles of intracellular regions in the activation of voltage-dependent potassium channels.
Eur. Biophys. J.,
2004
May
, 33 (194-200).
Yellen G
The voltage-gated potassium channels and their relatives.
Nature,
2002
Sep
5
, 419 (35-42).
Lörinczi E
et al.
The voltage dependence of hEag currents is not determined solely by membrane-spanning domains.
Eur. Biophys. J.,
2009
Mar
, 38 (279-84).
Zhang X
et al.
Divalent cations slow activation of EAG family K+ channels through direct binding to S4.
Biophys. J.,
2009
Jul
8
, 97 (110-20).
Jegla TJ
et al.
Evolution of the human ion channel set.
Comb. Chem. High Throughput Screen.,
2009
Jan
, 12 (2-23).
Bannister JP
et al.
Optical detection of rate-determining ion-modulated conformational changes of the ether-à-go-go K+ channel voltage sensor.
Proc. Natl. Acad. Sci. U.S.A.,
2005
Dec
20
, 102 (18718-23).
Tang CY
et al.
Extracellular Mg(2+) modulates slow gating transitions and the opening of Drosophila ether-à-Go-Go potassium channels.
J. Gen. Physiol.,
2000
Mar
, 115 (319-38).
Terlau H
et al.
Extracellular Mg2+ regulates activation of rat eag potassium channel.
Pflugers Arch.,
1996
Jun
, 432 (301-12).
Silverman WR
et al.
Binding site in eag voltage sensor accommodates a variety of ions and is accessible in closed channel.
Biophys. J.,
2004
Nov
, 87 (3110-21).
Silverman WR
et al.
Mg(2+) modulates voltage-dependent activation in ether-à-go-go potassium channels by binding between transmembrane segments S2 and S3.
J. Gen. Physiol.,
2000
Nov
, 116 (663-78).
Schönherr R
et al.
Conformational switch between slow and fast gating modes: allosteric regulation of voltage sensor mobility in the EAG K+ channel.
Neuron,
2002
Aug
29
, 35 (935-49).
Ju M
et al.
Molecular identification and characterisation of the human eag2 potassium channel.
FEBS Lett.,
2002
Jul
31
, 524 (204-10).
Ludwig J
et al.
Cloning and functional expression of rat eag2, a new member of the ether-à-go-go family of potassium channels and comparison of its distribution with that of eag1.
Mol. Cell. Neurosci.,
2000
Jul
, 16 (59-70).
Garg V
et al.
Tuning of EAG K(+) channel inactivation: molecular determinants of amplification by mutations and a small molecule.
J. Gen. Physiol.,
2012
Sep
, 140 (307-24).
Yang Y
et al.
Multistate structural modeling and voltage-clamp analysis of epilepsy/autism mutation Kv10.2-R327H demonstrate the role of this residue in stabilizing the channel closed state.
J. Neurosci.,
2013
Oct
16
, 33 (16586-93).
de Oliveira RM
et al.
Eag1, Eag2, and SK3 potassium channel expression in the rat hippocampus after global transient brain ischemia.
J. Neurosci. Res.,
2012
Mar
, 90 (632-40).
Gómez-Varela D
et al.
Characterization of Eag1 channel lateral mobility in rat hippocampal cultures by single-particle-tracking with quantum dots.
PLoS ONE,
2010
, 5 (e8858).
Schönherr R
et al.
Functional distinction of human EAG1 and EAG2 potassium channels.
FEBS Lett.,
2002
Mar
13
, 514 (204-8).
Jeng CJ
et al.
Differential localization of rat Eag1 and Eag2 K+ channels in hippocampal neurons.
Neuroreport,
2005
Feb
28
, 16 (229-33).
Huang X
et al.
Voltage-gated potassium channel EAG2 controls mitotic entry and tumor growth in medulloblastoma via regulating cell volume dynamics.
Genes Dev.,
2012
Aug
15
, 26 (1780-96).
Camacho J
et al.
Cytoskeletal interactions determine the electrophysiological properties of human EAG potassium channels.
Pflugers Arch.,
2000
Dec
, 441 (167-74).
Terlau H
et al.
Amino terminal-dependent gating of the potassium channel rat eag is compensated by a mutation in the S4 segment.
J. Physiol. (Lond.),
1997
Aug
1
, 502 ( Pt 3) (537-43).
Contributors: Rajnish Ranjan, Katherine Johnston
To cite this page: [Contributors] Channelpedia https://channelpedia.epfl.ch/wikipages/34/ , accessed on 2026 Aug 01