Kv3.3
Description: potassium voltage gated channel, Shaw-related subfamily, member 3 Gene: Kcnc3 Alias: Kv3.3, kcnc3, Kcr2-3, KShIIID
Kv3.3, encoded by the gene KCNC3, is a voltage-gated potassium channel. Kv3.3 plays a key role in high-frequency firing and motor coordination Mutations in Kv3.3 lead to disruptions in sound localization and contributing to alcohol hypersensitivity and spontaneous myoclonus.
Experimental data
Rat Kv3.3 gene in CHO host cells datasheet |
||
|
Click for details 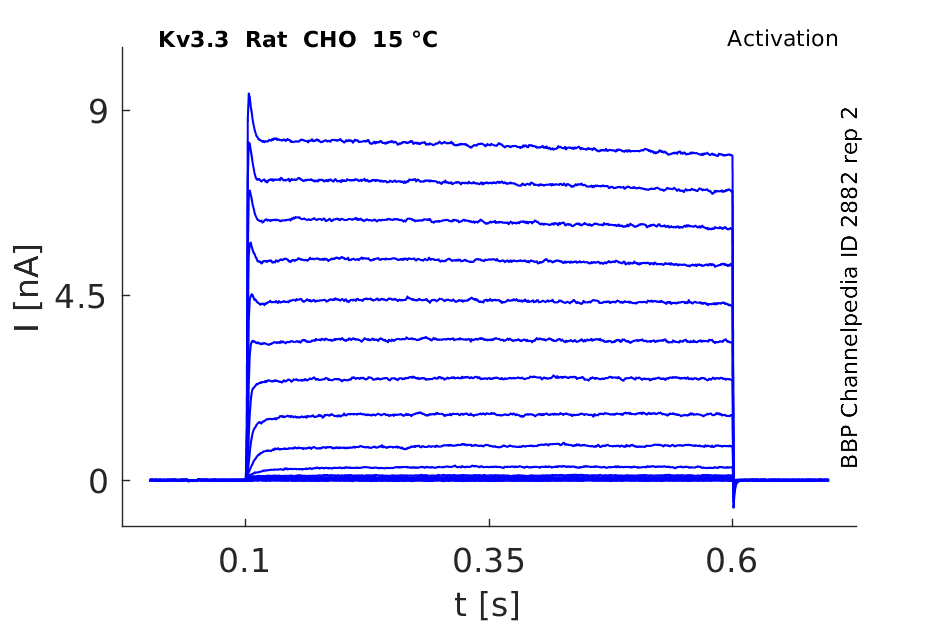
15 °Cshow 78 cells |
Click for details 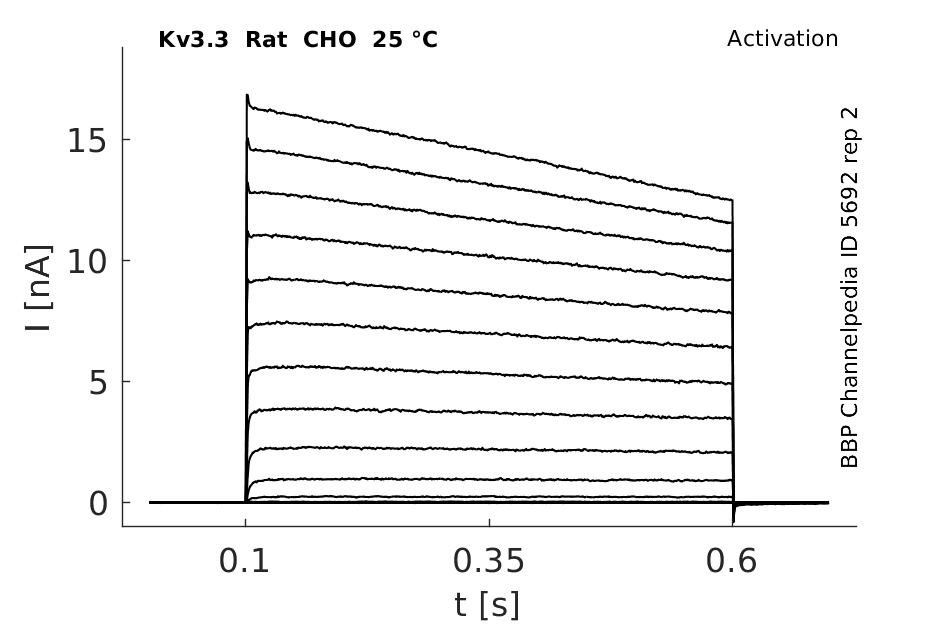
25 °Cshow 172 cells |
Click for details 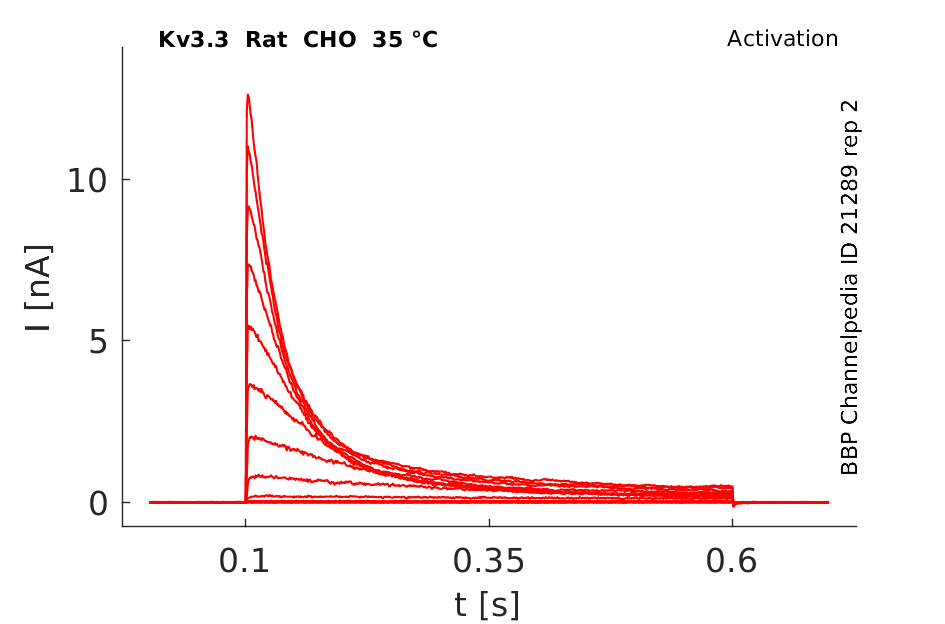
35 °Cshow 124 cells |
Gene
Transcript
| Species | NCBI accession | Length (nt) | |
|---|---|---|---|
| Human | NM_004977.3 | 6747 | |
| Mouse | NM_008422.3 | 5490 | |
| Rat | NM_053997.5 | 5268 |
Isoforms
Protein Kinase C (PKC)
Kv3.3 currents are potently enhanced by activation of PKC. Two N-terminal serine residues are consensus sites for PKC phosphorylation and are required for enhancement of Kv3.3 currents. PKC modulation of Kv3.3 channels could play a role in dynamic adaptation of auditory brainstem circuits. [20]
Kv3.3 predicted AlphaFold size
Methodology for AlphaFold size prediction and disclaimer are available here
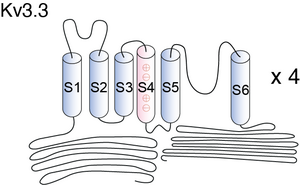
Visual Representation of Kv3.3 Structure
Methodology for visual representation of structure available here
Both Kv3.3b and Kv3.4 express as a transient or A-type current, but inactivation of Kv3.4 channels (τ = 15.9 ms) is much faster than Kv3.3 (τ = 200 ms) [1638]
SINGLE CHANNEL CONDUCTANCE


The single channel slope conductance measured under presumed equimolar K+ in the on-cell configuration ranged between 32 and 38 pS and was not significantly different for channels expressed in CHO or HEK cells. Single channel recordings were recorded in the on-cell or outside-out configuration and were acquired at 2–5 kHz and digitally filtered during analysis at 1–2 kHz. [1641]
Kv3.3 Channel Kinetics in CHO and HEK cells


Deactivation was rapid in both CHO and HEK cells (CHO τ = 0.55 to 1.09 ms; HEK τ = 0.41 to 0.81 ms) and voltage-dependent between –80 and –30 mV. As found for activation, the τ of deactivation in CHO cells was significantly slower than that in HEK cells [1641] Kv3.3 and Kv3.4 currents are of the A-type, although Kv3.3 currents inactivate slowly whereas Kv3.4 currents inactivate relatively quickly [302]
Biophysics
Model Kv3.3 (ID=28)
| Animal | Xenopus | |
| CellType | oocyte | |
| Age | 0 Days | |
| Temperature | 23.0°C | |
| Reversal | -65.0 mV | |
| Ion | K + | |
| Ligand ion | ||
| Reference | [280] A J Rashid et. al; J. Neurosci. 2001 Jan 1 | |
| mpower | 1.0 | |
| m Inf | 1/(1+exp(((v -(18.700))/(-9.700)))) | |
| m Tau | 20.000/(1+exp(((v -(-46.560))/(-44.140)))) | |

Model Kv3.3 (ID=45)
| Animal | CHO | |
| CellType | CHO | |
| Age | 0 Days | |
| Temperature | 23.0°C | |
| Reversal | 82.0 mV | |
| Ion | K + | |
| Ligand ion | ||
| Reference | [20] Leonard K Kaczmarek et. al; J. Biol. Chem. 2008 Aug 8 | |
| mpower | 2.0 | |
| m Inf | 1/(1+exp((v-35)/-7.3)) | |
| m Tau | 0.676808 +( 27.913114 / (1 + exp((v - 22.414149)/9.704638))) | |
| hpower | 1.0 | |
| h Inf | 0.25+( 0.75 /(1+exp((v-(-28.293856))/29.385636))) | |
| h Tau | 199.786728 + (2776.119438 * exp(-v/7.309565)) | |

Kv3.3 Expression in CNS

Kv3.3 is widely expressed in the nervous system, including cerebellum, basal ganglia and spinal cord [487], areas implicated in the precise execution of motor tasks. Kv3.3 is the dominant Kv3 family member in the auditory brainstem, for example in the calyces of Held, the large presynaptic terminals that provide input to the MNTB, and also in the superior olivary complex, where the axons of MNTB neurons end.[486]
In Purkinje cells, Kv3.3 is expressed in the absence of Kv3.1. [318]
Kv3.1 and Kv3.3 mRNA transcripts overlap in many areas, particularly in the posterior part of the brain and in the spinal cord. Kv3.3 are expressed in many neuronal populations, including most auditory central processing neurons (some of which also express Kv3.2) and many cranial nerve nuclei [302]
In the apteronotid electrosensory lateral line lobe, a homolog of the Kv3.3 K+ channel subtype is distributed over the entire soma–dendritic axis of pyramidal cells. [280]
Kv3.3 is distributed an axonal membranes and synaptic terminals where they influence spike repolarization [1639]
Mammalian Kv3.3 potassium channel, AptKv3.3, is expressed at high levels in both the somas and dendrites of ELL pyramidal cells. In addition, preliminary in situ hybridization studies indicated that a second Kv3 subtype, AptKv3.1, was also expressed in these neurons [1667]
Kv3.3-immunoreactivity was widespread in the vestibular nuclei and was detected in somata, dendrites and synaptic terminals
Kv3.3 in Dendrites
Voltage-gated potassium channel subunit Kv3.3 is expressed in the distal dendrites of Purkinje cells. However, the functional relevance of this dendritic distribution is not understood. To study the physiological relevance of altered dendritic excitability, we measured Ca(2+) changes throughout the dendritic tree in response to climbing fiber activation. Ca(2+) signals were specifically enhanced in distal dendrites of Kv3.3 knockout Purkinje cells, suggesting a role for dendritic Kv3.3 channels in regulating propagation of electrical activity and Ca(2+) influx in distal dendrites [1666]
NEURONAL FUNCTION
Due to their biophysical properties, Kv3 channels facilitate firing of narrow action potentials at high frequency, and their interaction with resurgent Na+ currents support spontaneous action potential firing in the absence of synaptic inputs. In a previous electrophysiological study, it was shown that neurons of deep cerebellar nuclei (DCN) lacking glutamic acid decarboxylase (GAD-), the GABA-synthesizing enzyme, fire at higher rates and exhibit faster action potentials than GAD+ DCN neurons. These observations correlated with a distinct distribution of Kv3.1b and Kv3.3 potassium channel subunits in GAD- and GAD+ dentate neurons [1639]
The functional properties of zebrafish Kv3.3 channels are consistent with a role in facilitating fast, repetitive firing of action potentials in neurons. The functional effects of SCA13 mutations are well conserved between human and zebrafish Kv3.3 channels [1664]
KO MICE BEHAVIOUR
Mice lacking both Kv3.1 and Kv3.3 subunits display severe motor disturbances, such as ataxia, tremor, myoc- lonus as well as high alcohol hypersensitivity, while there is only a modest motor dysfunction in mice lacking only one of the subunits (Kv3.1 or Kv3.3). Indeed, the number of intact Kv3.1/3.3 alleles correlates with the severity of ataxia, indicating some degree of functional redundancy at least in granule cell parallel fibers [1639]
In the apteronotid electrosensory lateral line lobe, a Kv3.3 K+ channel subtype acts to repolarize Na+ spike discharge in both the soma and proximal apical dendrites. [280]
Mice lacking either Kv3.1 or Kv3.3 subunits display relatively moderate electrophysiological and behavioral changes [489]. Knocking out both, Kv3.1 and Kv3.3, however, ensues dramatic physiological and behavioral alterations that include hyperactivity, sleep loss, myoclonus, tremor, alcohol hypersensitivity and severe ataxia [488].
Sound Localization
Mutation in the Kv3.3 Voltage-Gated Potassium Channel Causing Spinocerebellar Ataxia 13 Disrupts Sound-Localization Mechanisms
Spinocerebellar Ataxia
Mutations in Kv3.3 cause the neurological disorder SCA13 (spinocerebellar ataxia type 13) [1665]
Alcohol Hypersensitivity
Alcohol hypersensitivity, increased locomotion, and spontaneous myoclonus in mice lacking the potassium channels Kv3.1 and Kv3.3 [488]
TEA

In both cell types, substantial block was obtained for whole-cell currents in the range of 200 μM TEA or less. There was a significantly lower IC50 for whole-cell currents expressed in CHO compared with HEK cells (CHO IC50 = 67.5 ± 4.2 μM, HEK IC50 = 152 ± 27 μM). The IC50 value for TEA obtained in HEK cells was equivalent to that reported for Kv3.3a expressed in oocytes at 140 ± 43 μM. AptKv3.3 currents expressed in HEK cells were also highly sensitive to externally applied 4-AP (IC50 < 100 μM) [1641]
Mutations
Zebrafish Kv3.3 activates over a depolarized voltage range and deactivates rapidly. An amino-terminal extension mediates fast, N-type inactivation. The kcnc3a gene is alternatively spliced, generating variant carboxyl-terminal sequences. The R335H mutation in the S4 transmembrane segment, analogous to the SCA13 mutation R420H, eliminates functional expression. When co-expressed with wild type, R335H subunits suppress Kv3.3 activity by a dominant negative mechanism. The F363L mutation in the S5 transmembrane segment, analogous to the SCA13 mutation F448L, alters channel gating. F363L shifts the voltage range for activation in the hyperpolarized direction and dramatically slows deactivation [1664]
References
Desai R
et al.
Protein kinase C modulates inactivation of Kv3.3 channels.
J. Biol. Chem.,
2008
Aug
8
, 283 (22283-94).
Rashid AJ
et al.
The contribution of dendritic Kv3 K+ channels to burst threshold in a sensory neuron.
J. Neurosci.,
2001
Jan
1
, 21 (125-35).
Rudy B
et al.
Kv3 channels: voltage-gated K+ channels designed for high-frequency repetitive firing.
Trends Neurosci.,
2001
Sep
, 24 (517-26).
Joho RH
et al.
Behavioral motor dysfunction in Kv3-type potassium channel-deficient mice.
Genes Brain Behav.,
2006
Aug
, 5 (472-82).
Joho RH
et al.
Kv3 potassium channels control the duration of different arousal states by distinct stochastic and clock-like mechanisms.
Eur. J. Neurosci.,
2006
Mar
, 23 (1567-74).
Chang SY
et al.
Distribution of Kv3.3 potassium channel subunits in distinct neuronal populations of mouse brain.
J. Comp. Neurol.,
2007
Jun
20
, 502 (953-72).
Kopp-Scheinpflug C
et al.
The medial nucleus of the trapezoid body in the gerbil is more than a relay: comparison of pre- and postsynaptic activity.
J. Assoc. Res. Otolaryngol.,
2003
Mar
, 4 (1-23).
Li W
et al.
Localization of two high-threshold potassium channel subunits in the rat central auditory system.
J. Comp. Neurol.,
2001
Aug
20
, 437 (196-218).
Brooke RE
et al.
Spinal cord interneurones labelled transneuronally from the adrenal gland by a GFP-herpes virus construct contain the potassium channel subunit Kv3.1b.
,
2002
Jun
28
, 98 (45-50).
Espinosa F
et al.
Alcohol hypersensitivity, increased locomotion, and spontaneous myoclonus in mice lacking the potassium channels Kv3.1 and Kv3.3.
J. Neurosci.,
2001
Sep
1
, 21 (6657-65).
Ho CS
et al.
Pleiotropic effects of a disrupted K+ channel gene: reduced body weight, impaired motor skill and muscle contraction, but no seizures.
Proc. Natl. Acad. Sci. U.S.A.,
1997
Feb
18
, 94 (1533-8).
Minassian NA
et al.
Altered Kv3.3 channel gating in early-onset spinocerebellar ataxia type 13.
J. Physiol. (Lond.),
2012
Apr
1
, 590 (1599-614).
Veys K
et al.
Kv3.3b expression defines the shape of the complex spike in the Purkinje cell.
Front Cell Neurosci,
2013
, 7 (205).
Puente N
et al.
Precise localization of the voltage-gated potassium channel subunits Kv3.1b and Kv3.3 revealed in the molecular layer of the rat cerebellar cortex by a pre-embedding immunogold method.
Histochem. Cell Biol.,
2010
Oct
, 134 (403-9).
Brooke RE
et al.
Kv3.3 immunoreactivity in the vestibular nuclear complex of the rat with focus on the medial vestibular nucleus: targeting of Kv3.3 neurones by terminals positive for vesicular glutamate transporter 1.
Brain Res.,
2010
Jul
23
, 1345 (45-58).
Fernandez FR
et al.
Inactivation of Kv3.3 potassium channels in heterologous expression systems.
J. Biol. Chem.,
2003
Oct
17
, 278 (40890-8).
Sung MJ
et al.
Effect of psoralen on the cloned Kv3.1 currents.
Arch. Pharm. Res.,
2009
Mar
, 32 (407-12).
Mock AF
et al.
Functional effects of spinocerebellar ataxia type 13 mutations are conserved in zebrafish Kv3.3 channels.
,
2010
Aug
16
, 11 (99).
Zhao J
et al.
Spinocerebellar ataxia-13 Kv3.3 potassium channels: arginine-to-histidine mutations affect both functional and protein expression on the cell surface.
Biochem. J.,
2013
Sep
1
, 454 (259-65).
Zagha E
et al.
Dendritic Kv3.3 potassium channels in cerebellar purkinje cells regulate generation and spatial dynamics of dendritic Ca2+ spikes.
J. Neurophysiol.,
2010
Jun
, 103 (3516-25).
Deng Q
et al.
A C-terminal domain directs Kv3.3 channels to dendrites.
J. Neurosci.,
2005
Dec
14
, 25 (11531-41).
Rae JL
et al.
Kv3.3 potassium channels in lens epithelium and corneal endothelium.
Exp. Eye Res.,
2000
Mar
, 70 (339-48).
Liu SQ
et al.
Aminoglycosides block the Kv3.1 potassium channel and reduce the ability of inferior colliculus neurons to fire at high frequencies.
J. Neurobiol.,
2005
Mar
, 62 (439-52).
Contributors: Rajnish Ranjan, Michael Schartner, Nitin Khanna, Katherine Johnston
To cite this page: [Contributors] Channelpedia https://channelpedia.epfl.ch/wikipages/13/ , accessed on 2026 Jul 18